Abstracts
On the occasion of having collected three gravid Geochelone carbonaria in the Bolivian Chaco, data are given on oviducal eggs, as well as on five shells of eggs of the same species in the Museum collection. A review of the literature on the reproduction of G. carbonaria and of its sibling G. denticulata is presented, with an elementary statistical treatment of the data on number of clutches per reproductive season, interval between clutches, clutch size, egg shape and volume.
Testudines; reproduction
São apresentadas notas a respeito de ovos oviducais de três exemplares de G. carbonaria, coletados no Chaco boliviano, bem como a respeito de cinco cascas de ovos da coleção do Museu. É também apresentada uma revisão da literatura a respeito de reprodução de G. carbonaria e da espécie irmã G. denticulata, com um tratamento estatístico elementar dos dados a respeito do número de posturas por ano, intervalo entre posturas, tamanho da ninhada, forma e volume dos ovos.
Testudines; reprodução
A NOTE ON THE REPRODUCTION OF geochelone carbonaria AND G. denticulata (TESTUDINES, TESTUDINIDAE)
VANZOLINI, P. E.
Museu de Zoologia, Universidade de São Paulo, C.P. 42694, CEP 04299-970, São Paulo, SP, Brazil
Correspondence to: Paulo Emilio Vanzolini, Museu de Zoologia, Universidade de São Paulo, C.P. 42694,
CEP 04299-970, São Paulo, SP, Brazil
Received August 10, 1998 Accepted September 26, 1998 Distributed December 22, 1999
(With 4 figures)
ABSTRACT
On the occasion of having collected three gravid Geochelone carbonaria in the Bolivian Chaco, data are given on oviducal eggs, as well as on five shells of eggs of the same species in the Museum collection. A review of the literature on the reproduction of G. carbonaria and of its sibling G. denticulata is presented, with an elementary statistical treatment of the data on number of clutches per reproductive season, interval between clutches, clutch size, egg shape and volume.
Key words: Testudines, reproduction.
RESUMO
Nota sobre a reprodução de Geochelone carbonaria e G. denticulata (Testudines, Testudinidae)
São apresentadas notas a respeito de ovos oviducais de três exemplares de G. carbonaria, coletados no Chaco boliviano, bem como a respeito de cinco cascas de ovos da coleção do Museu. É também apresentada uma revisão da literatura a respeito de reprodução de G. carbonaria e da espécie irmã G. denticulata, com um tratamento estatístico elementar dos dados a respeito do número de posturas por ano, intervalo entre posturas, tamanho da ninhada, forma e volume dos ovos.
Palavras-chave: Testudines, reprodução.
INTRODUCTION
The tortoises of the genus Geochelone, and especially the widely distributed G. denticulata (L., 1766) and G. carbonaria (Spix, 1824), are common in their area of occurrence. They are docile and well-adapted captives, and captivity in no way abates their considerable amatory proclivities. It is surprising how little is known of their reproductive biology, especially in the wild.
On November 17, 1997, a Museum party (Celso Morato de Carvalho, Fernando Mendonça d'Horta and myself), travelling through the Bolivian Chaco, collected four adult specimens, one male and three females, of G. carbonaria (catalogued as MZUSP 4002-4005), on a stretch of road 13 km long south of Fortin Suarez Arana, on the way to Fortin Ravelo, in the province of Santa Cruz. The approximate coordinates of the extreme points of collection are 18°56'S, 60°20'W, and 19°01'S, 60°24'W. Not having drums large enough to preserve the specimens in fluid, we followed our procedure for such cases, dissecting the head, neck, limbs and tail, to be preserved in formalin, sawing through the bridges, scraping and drying the shell. In the process it was noticed that the three females were gravid.
Given the dearth of information on the species in general, and on Chaco populations specifically, and in accordance with the principle that the killing of a wild animal is an ethically serious act, demanding that maximum scientific profit be extracted from the kill, I think it desirable to present the data, as obtained in the field, complemented with additional information from our collection, and, of course, a review of the literature, which is rather unsatisfactory. In fact, the main reviews (in Harless & Morlock, 1979; in Gans & Huey, 1988) provide very meager and generalized information.
Disembowelling turtles in the middle of nowhere for taxidermical purposes is of course not an ideal way of acquiring scientific information. Our materials, however, have potential interest regarding some aspects of reproductive biology. It is also clear that a review of the literature, scattered, uneven and at times hard to locate, is overdue.
LITERATURE
In order to build a frame of reference on the reproductive biology of South American Geochelone, portraying the state of the art and the more obvious needs for further information, I have made an effort to bring together and organize the available data on the aspects relevant to the present matter, not only of Geochelone carbonaria, but also of G. denticulata. They are true siblings and extensively sympatric (Auffenberg, 1971; Williams, 1960); it is worthwhile to consider them together. As said, I have limited myself to the aspects of reproductive biology to which the present materials are relevant: courtship, nesting, incubation and fertility are not treated.
The literature on other species of Geochelone, South American or not, has also been searched, although it will not be cited here and only discussed, when pertinent, under "Comments". I have entirely omitted the literature on Gopherus, because this taxon is phylogenetically and ecologically too distant to be included in the context of this report.
The earliest references to reproductive aspects of South American Geochelone are in Bell's (1836-1842) texts accompanying the plates depicting Testudo tabulata (= G. denticulata), in part 1, and T. carbonaria, in part 2 of his "Monograph of the Testudinata"; these parts were published in 1836 (see "References" below). For denticulata he says: "The eggs are of a spherical shape, very slightly flattened, and about two inches in diameter. The egg shell is thick and of a friable texture". For G . carbonaria: "The egg is very similar to that of T. tabulata, but rather smaller, being an oblate spheroid, of which the greater diameter is 1 inch 9 lines and the less 1 inch 6 lines".
The next author on Geochelone biology is Major João Martins da Silva Coutinho, travel companion and factotum of Louis Agassiz during the latter's Amazonian expedition, 1865-1866 (Agassiz & Agassiz, 1868: 122; Vanzolini, 1996). The Major, although a career soldier (corps of engineers), was a cultured man and had a fine feeling for natural history. He published in Europe (see Vanzolini, 1977) two papers, in 1868 and 1886, on Amazonian chelonians. In the 1868 paper, apparently written at the behest of Duméril fils, he mentions (Coutinho, 1868: 156) three species of Geochelone, using for them names from the Tupí Indian language, widespread, usual and traditional in Brasil: (i) "Jaboty-tinga", which he identified as Testudo carbonaria; (ii) "Jaboty-piranga", no scientific name offered; and (iii) "Jaboty-carumbé", also not identified. "Jabotí" was then , and continues to be, the Tupí name, universal in Brasil, for Geochelone.
In a footnote to Coutinho's paper, Auguste Duméril comments on the identifications. He correctly notes that Coutinho mixed up the diagnostic characters of his tortoises. "Tinga" (Tupí, white tortoise) would have, according to the Major, a constricted carapace and yellow spots on the head and feet. "Piranga" (Tupí, red tortoise) would have red spots and no constriction in the carapace. The real association is the reverse: G. carbonaria, the red foot, has a constricted shell and red spots; G. denticulata, the yellow foot, has a rounded shell and yellow spots. As to "Jaboty-carumbé", Duméril could offer no guesses. It is understandable: "carumbé" is simply the Amazonian Tupí word for male tortoises in general (Goeldi, 1906: 713; Tastevin, 1923: 705). The term is still current in Amazonia.
Coutinho (1868: 155) states that the three species had identical breeding habits, and that all laid spherical eggs. In the specific case of jaboty tinga he adds that the clutch comprises ten to fifteen eggs, deposited in a nest covered by the female. I think, with Duméril (loc. cit.), that, in spite of the confusion about the color pattern, Coutinho's identification stands, and the data may be attributed to G. carbonaria, keeping always in mind that it is nowhere stated how the Major came by his information, if by personal observation (the animals are common), or by hearsay (they are well known).
Ihering (1904), reporting on a general zoological collection made by Ernesto Garbe in the Rio Juruá, state of Amazonas, mentions, for "Testudo tabulata", two eggs, almost globular in shape, with a hard calcareous shell. The tortoise is in our collection and is G. denticulata; the eggs (measurements of which are given by Ihering) are not to be found. Francisco Dias da Rocha (1911: 40) cites measurements of the eggs of "Testudo tabulata". Rocha lived in the state of Ceará, so the tortoise is probably G. carbonaria (Williams, 1960).
Snedigar & Rokoski (1950) mention two clutches of G. denticulata, no locality given, in the Chicago (Brookfield) zoo: they give measurements and weights of individual eggs. Medem (1960) reports for G. denticulata from Amazonian Colombia clutches of 5-15 "large" (averages 50 x 42 mm) eggs; Underwood (1962), on Antillean G. carbonaria from the Grenadines, with measurements of two eggs; Legler (1963) on G. carbonaria from the Panamanian provinces of Darién and San Blas, giving estimates of clutch size, from dissection.
Dahl & Medem (1964: 117) report on both G. denticulata and G. carbonaria. Although the paper is a joint one, the account is written in the first person singular, so it is safe to attribute it to Medem, who was a herpetologist much concerned with Colombian chelonians. There are notes on four clutches of denticulata in captivity. Measurements are given, and the weight of one egg. On G. carbonaria Medem's information stems from a private breeder. Medem's data stand out in the meagre literature, and are often cited. I am, however, in favor of altogether disregarding them: there is an assertion that clutches are always in odd numbers, 1, 3 or 5, never 2 or 4. To me, such arrant idiosyncrasy compromises the whole set of data. It is to be noted that Medem's data are not always easy to evaluate. Fortunately this research was followed by two of his students, Olga Victoria Castaño-Mora and Myriam Lugo-Rugeles (see below), and excellent data are available for Amazonian Colombia.
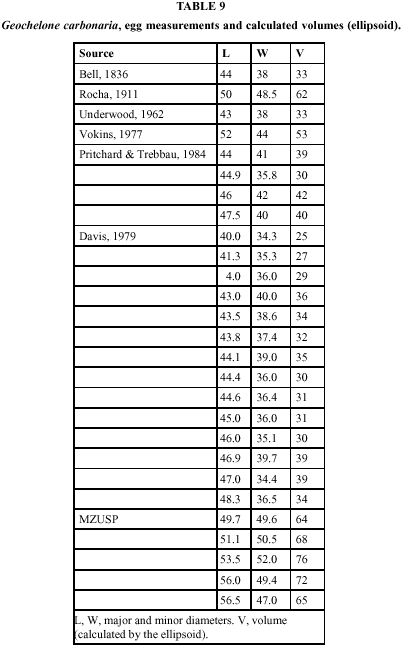
Vokins (1977) has data on the number of eggs in fourteen clutches of G. carbonaria laid in the Jersey zoo. He also gives average measurements and weights of eggs for one of the clutches. Medem, Castaño-M. and Lugo-R. (1979), give succint data on clutch size and egg measurements for G. carbonaria, apparently kept in captivity at their laboratory in Villavicencio (see below). Moll (1979), in a review article, places in a bar diagram, without specific references, G. carbonaria in a class having the "mean or usual clutch size" of 8-10 eggs, G. denticulata in the 15 eggs class. Davis (1979) gives data on G. carbonaria reproducing in the Washington zoo; of interest are measurements and weights of individual eggs.
By far the best set of data available in the literature is that of Castaño-Mora and Lugo-Rugeles (1981). Although the title of the paper does not mention reproduction, it contains a wealth of data. They worked with the herd already mentioned, kept by their former mentor, Federico Medem, at the Instituto Roberto Franco, Villavicencio, Colombia. Both species are included and data given on number of clutches, on clutch size and (practically unique in the literature) interval between successive clutches. The carbonaria series is excellent, the denticulata one is smaller. The observations were carried over 1977 and 1978. Only raw data are presented; I have submitted them (below) to some elementary statistical treatment.
Pritchard & Trebbau (1984) present incidental data on clutch size and egg measurements for both species. Moskovits (1985) has also incidental data on clutch size of G. carbonaria. Rocha et al. (1988), in an abstract of a congress presentation, show, for G. carbonaria in a zoo, among other data, ranges and means of clutch size and of egg weight. Guix et al. (1989) give data on clutch size of G. carbonaria in captivity. Tonge (1988) extends Vokins's (1977) data. Moreira (1991) presents data on five gravid G. denticulata from the middle Amazon. Her data are incomplete and restricted to ranges, but they are the only so far obtained from wild specimens.
Mallmann (1994), in an unpublished master's dissertation on the influence of incubation temperature on sex determination, gives incidental data on clutch size and egg measurements.
DATA
The Chaco specimens
The largest specimen (MZUSP 4005, plastral length 285 mm) had eggs on the left side only. Ten eggs were large, not shelled, consequently spherical. Their diameters (taken in the field with a straight ruler) and weights (taken at the same time with Pesola scales) are shown on Table 14. Fifteen small eggs, diameter 8-10 mm, were not measured, just weighed jointly (3.8 g.). There were additionally numerous large ovarian follicles. The smallest female (MZUSP 4003, plastral length 240 mm) had (also unshelled) eggs only on the left side: the measurements and weights of three large eggs are discused below; there were additionally eleven eggs 11-16 mm in diameter, not weighed; also follicles. The third female, intermediate in size (MZUSP 4004, plastral length 270 mm), had (again unshelled) eggs on both sides. On the left side it had two large (Table 1) and seven small eggs, the latter measuring 16-23 mm in diameter and weighing jointly 23 g. On the right side there were nine eggs, a few scattered small ones and many enlarged follicles.

Other materials
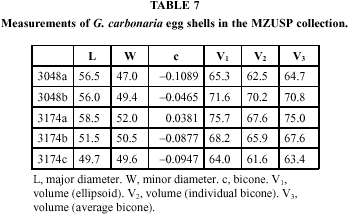
Besides this new information, we have in our collection two samples of Geochelone carbonaria egg shells: one sample of two eggs (MZUSP 3048) from a zoo specimen, one of three eggs (MZUSP 3174) from a female collected in the area of Aquidauana, Mato Grosso do Sul, contributed by Arlindo Béda (Table 2, Figs. 1, 2), to whom thanks are due. Our materials, taken together, have potential interest regarding five aspects of reproductive biology: (i) timing of reproduction; (ii) clutch size; (iii) number of clutches per season; (iv) weight of unshelled eggs; (v) size and shape of laid eggs.
COMMENTS
Number of clutches per season (Table 1)
Besides the definite data of Castaño & Lugo (1981) and of Tonge (1988), and the assertion of Rocha et al. (1988) of at least two clutches per year, there is much indirect evidence for multiple clutches in G. carbonaria, principally the simultaneous presence of eggs at different stages of maturation.
The present data belong in this class: either one or both oviducts occupied, oviducal and ovarian eggs present in the same female.
Given the large number of successive clutches (up to 5) observed by the authors cited, it is clear that autopsy data do not permit an estimation of the number of clutches in one reproductive season.
Castaño & Lugo's (1981) and Tonge's (1988), data are shown on Table 1. The former, obtained in the tortoises' homeland, in semi-natural conditions, may be taken as tentative paradigms for the species and region, not forgetting that the G. carbonaria sample is by far the best. McKeown, Juvik & Meier (1982) cite two clutches per year, irregularly spaced, for the Madagascar species G. yniphora in the Honolulu Zoo.
Interval between clutches (Tables 2 and 3)
Again I refer to Castaño & Lugo (1981). They give the actual dates of laying, and the respective intervals, for the years 1977 and 1978. (My reckonings differ topically from theirs by one day; I am using my own counts). It seems to me that the most salient point in Tables 2 and 3 is the very large spread of the data, as indicated by the ranges and coefficients of variation.
There are no significant differences between the intervals between successive clutches of G. carbonaria. The interval between first and second clutches does not differ significantly between the two species (Mann-Whitney test, p > 0.90).
Tonge (1988) has the data of a 20-year longitudinal study of a pair of G. carbonaria at the Jersey zoo. The data agree well with those of Castaño & Lugo (1981); they contain the additional information that no eggs were laid in four different years there was no clutch laid.
Clutch size (Tables 4 to 6)
Table 4 lists the data on G. carbonaria in the literature. It shows remarkable spread both within and between observers. Table 5 shows the statistics of the cases where the published evidence includes appropriate raw data. Again the very high coefficients of variation should be noted.
Castaño & Lugo's (1981) data permit another interesting piece of analysis (Table 6): there is no statistically significant variation in the number of eggs in successive clutches.
Finally, it is to be seen that the average number of eggs per clutch is larger in G. denticulata (t = 5.154, p < 0.1%).
Galapagoan G. elephantopus is known to lay 7-22 eggs per clutch (Hairston & Burchfield, 1989); Burmese G. emys, 23-51; and Madagascan G. yniphora 3-6 (Mc Keown et al., 1982); all in zoos.
Shape and volume of the eggs (Tables 7 and 8)
Volume. The ascertainment of the geometrical form of the egg is essential to the calculation of the volume, whose direct measurement is usually difficult and uncertain (Preston, 1953); in the present case it was impracticable by any unsophisticated means. Additionally, indirect estimation is necessary for the bare linear measurements in the literature.
The current approach to the indirect estimation of egg volume originated in the seminal paper by Preston (1953) on birds' eggs. It was extended to reptiles by Iverson & Ewert (1991) and, principally, by Maritz & Douglas (1994), whose methodology was adopted by subsequent authors (Rose et al., 1996; Vanzolini, 1997).
Using adequate photographs (or scanned images) of individual eggs it is extremely easy to calculate, by a method of Maritz & Douglas (1994), the "bicone" of the egg, a measure of its departure from an ideal ellipsoid of revolution (prolate spheroid) having the same diameters. The concept of bicone originated with Preston (1953), who denoted it by c2 (c in the present paper). If the departure is considered not significant (no test of significance seems to have been devised), one may apply the formula for the volume of the ellipsoid
Ve =LW 2/6000
in which L and W are the "length" and "width" (major and minor diameters), measured in millimeters, which results in the volume being given in cubic centimeters. If the bicone is considered significant, one should apply a formula of Maritz & Douglas (1994)
V = Ve ((3c2 + 14 c + 35)/35)
Rose, Simpson & Manning (1996) suggest that c may be taken as the value for the individual egg, or, alternatively, as an average for the species (which latter alternative they favor). I calculated (Tables 7 and 8) the bicones of the 5 shells in our collection, and the volumes by the three methods outlined: ellipsoid, individual bicone and average (for the 5 specimens) bicone. It can be seen that the average of the bicones (as could be predicted from the very wide ranges) does not differ significantly from 0 (t = 2.252, p > 0.05); this, incidentally, sidesteps the need for a significance test of c. The three estimates of volume agree among themselves (analysis of variance, F = 0.106, not significant at the 5% level). In consequence, I feel justified in calculating, for present purposes, the volume of eggs of which diameters only are given in the literature, by the formula for the ellipsoid.
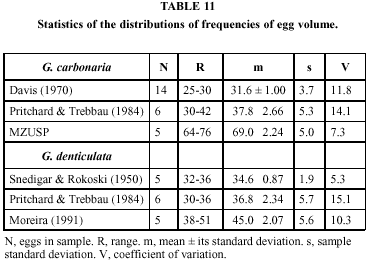
Tables 9 and 10 show the volumes thus calculated for the shells in our collection and for all eggs in the literature for which diameters are published. Table 11 summarizes the respective statistics. The distributions have reasonable coefficients of variation, especially considering that the volumes computed are products of two measurements which have their own variability.
Analysis of variance shows that the samples of G. carbonaria are a heterogeneous ensemble (F = 142.164, p < 0.001). Tukey's test, applied in sequence, shows that no two samples agree between themselves. In the case of G. denticulata the contrasts are relatively less sharp (F = 8.121; even so, p < 0.01); the means of Snedigar & Rososki and of Pritchard & Trebbau (1984) agree between themselves and disagree with Moreira's (1991).
The rationale behind these discrepancies lies probably in the field of geographic differentiation, but present materials are insufficient to judge.
This comprehensive heterogeneity makes it difficult (perhaps even senseless) to compare the two species. If this is to be attempted, though, there are two ways: (i) comparing directly Pritchard & Trebbau's samples of the two species, or (ii) doing an analysis of variance, followed by Tukey's test, for all samples of both species taken together.
The two samples of Pritchard & Trebbau closely agree (t = 0.295, p > 0.70). As to the results of the analysis of variance, the results are as follows (denoting G. carbonaria as C and G. denticulata as D, and indicating by a continuous line the samples that do not differ significantly):

i.e., the MZUSP and Moreira samples stand out, the volumes of our (MZUSP) eggs being exceedingy larger than the remainder. In fact, they are comparable to the eggs of Geochelone gigantea, a much larger species, that reaches 65 cm³ (Deraniyagala, 1960).
No systematic difference can thus be found between the two species in what regards egg volume, calculated as here done.
Shape. The numerical description of the form of the eggs has its own importance, besides the indirect calculation of the volume. In the present case this importance lies principally in the comparison of the two species.
Since it is legitimate to consider the egg as an ellipsoid of revolution, the geometrical properties of the solid must serve as a basis for analysis. There are two main ways of evaluating the departure of an ellipsoid from the sphere: the ratio of the two diameters, and the eccentricity of the generating ellipse.
A preliminary analysis showed that in no case is there significant regression between the two diameters; their ratio is thus compromised. The eccentricity is given by
e = SQR ((L² W²)/2L)
I calculated the eccentricities for all eggs for which diameters are available, but present here only the data for the samples with 4 or more eggs (Tables 12 and 13).
It is once more seen that there is very large variation, within and among species. Applying the same strategy as done for the volume:
Analysis of variance shows that both species are heterogeneous with regard to egg eccentricity (carbonaria, F = 4.848, p < 0.05; denticulata, F = 26.554, p < 0.001). Tukey's test shows no regularities for either species. Pritchard & Trebbau's (1984) two samples do not differ significantly (t = 1.616, p > 0.10). Joint analysis of variance followed by Tukey's test gives the following result:

No clear interspecific difference is apparent.
Timing of reproduction
The problem of the timing of reproduction in the case of animals with multiple clutches per year cannot be conclusively adressed by means of short term observations. Unless a wild population is followed for a significant period, isolated observations will have only sedimentary value. I now add two records of this type: the present one and that of Moreira (1991).
Calendrical dates by themselves are of course of no use; they should be correlated with environmental factors. In the case of the reproductive biology of tropical animals, the main environmental factor is rainfall.
I found on November 17 three females in full reproduction. I tried in vain to obtain climatological information on the Bolivian Chaco; the best I could do was data on the Paraguayan Chaco (Gorham, 1973). I selected from his data the two stations geographically closest to the area of interest: Bahia Negra (20º15'S, 58º12'W) and Mariscal Estigarribia (22º02'S, 60º38'W) it will be remembered that my place of collection was approximately at 18º 59'S, 60º22'W. In spite of the distance, ca. 300 km, the localities are ecologically very similar (I know them), and so I have adopted, as a first approximation, the Paraguayan data.
Moreira's (1991) specimens came for the basin of the Rio Uatumã, a tributary of the Amazonas on its left bank. The actual localities (Moreira, 1989) cluster around the mouth of a small stream, Igarapé Caititu, at approximately 01º44'S, 59º38'W.
The nearest meteorological station (Instituto Nacional de Meteorologia, 1979) is Manaus, on the same side of the Amazonas, at 03º07'S, 60º00'W, some 160 km to the SSW, but definitely within the same ecology (I also know the area).
It is to be seen on Fig. 1 that our specimens were in full reproductive activity a little (2 months) before the peak of the Chaco rainy season; Moreira's specimens were reproducing a similar two months but after the peak in Manaus. Without overstressing the significance of two isolated observations, the data suggest that reproduction in G. carbonaria is either linked to the rainy season, with a long reproductive period (in keeping with the presence of multiple clutches), or that the reproductive period is continuous, without a causal relationship to the pluviosity.
Oviducal eggs
I took in the field measurements and weights (mass) of the larger unshelled eggs. Diameter was measured with a straight ruler: this measurement of soft eggs cannot be accurate, even in replication. Weights, taken with adequate Pesola spring scales, can be accepted with security to the nearest gram. The raw data are shown on Table 14 and the statistics on Table 15. Analysis of variance shows that there is no significant heterogenety among the clutches (F = 1,144 for 2 and 21 degrees of freedom), so the females must have been well synchronized reproductively. An examination of graphs, linear and of the usual anamorphoses (arith-log, log-arith and log-log), indicated that the relationship between mass and volume can be described both by a linear regression and by a log-log one (logarithm of mass, on logarithm of volume) (Table 16, Fig. 2). Both models give, applied to all eggs taken together, equivalent fits, in fact good ones, as shown by the coefficients of determination, both in the neighborhood of 0.90.
These data are of course given for a descriptive purpose only.
Acknowledgments The trip through the Bolivian and Paraguayan Chaco was undertaken as part of an ecological-agricultural diagnosis (PRODEAGRO) of the Brasilian state of Mato Grosso, promoted by the state's Secretariat of Planning (SEPLAN), financed by the World Bank and contracted to the São Paulo firm of consultants "Consorcio Nacional de Engenheiros Consultores" (CNEC). W. R. Heyer, C. W. Myers, Esteban O. Lavilla and G. R. Zug kindly read the manuscript. Their contribution has been uniformly valuable, but as I have not uniformly accepted their suggestions, I want to stress that the shortcomings are mine. F. Val drew the graphs. For the climatic data of Paraguay I am indebted to W. R. Heyer and Mercedes S. Foster.
References
- AGASSIZ, L. & AGASSIZ, E. C., 1868, A journey in Brazil Boston: Ticknor & Fields. x + 540p.
- AUFFENBERG, W., 1969, Land of the Chaco tortoise. Int. Turtle Tort. Soc. J., 3(3): 16-19, 36-38.
- AUFFENBERG, W., 1971, A new fossil tortoise with remarks on the origin of South American testudinines. Copeia, 1971(1): 106-117.
- BELL, T., 1836-1842. A monograph of the Testudinata. Parts 1-8 London: Samuel Higley. xxiv + 82 p., pls. (Dates in Sherborn, 1922.)
- CASTAÑO-MORA, O. V. & LUGO-RUGELES, M., 1981, Estudio comparativo del comportamiento de dos especies de morrocoy: Geochelone carbonaria y Geochelone denticulata y aspectos comparables de su morfologia externa. Cespedesia, 10(37-38): 55-122.
- COUTINHO, J. M. S., 1868, Sur les tortues de l'Amazone. Bull. Soc. Imper. Zool. Acclim, 5(2): 147-166.
- COUTINHO, J. M. S., 1886, Bedeutung, Fang und Verwertung der Schildkröten am Amazonas. Zool. Gart., 27(11): 329-365, 366-372.
- DAHL, G. & MEDEM, F., 1964, Informe sobre la fauna acuatica del Rio Sinu Bogotá: Corporación Autónoma Regional de los Valles del Magdalena y del Sinu – CVM, 160p.
- DAVIS, S., 1979, Husbandry and breeding of the Red-footed tortoise, Geochelone carbonaria, at the National Zoological Park, Washington. Int. Zoo Yearb, 19: 50-53.
- DERANIYAGALA, P. E. P., 1960, Two giant tortoises in the Colombo National Museum. Spolia Zeyl., 29: 19-22.
- GANS, C. & HUEY, R. B., 1988, Biology of the Reptilia, vol. 16, Ecology B, Defense and life history New York, Alan R. Liss, xi + 659p.
- GOELDI, E. A., 1906, Chelonios do Brazil. Bol. Mus. Paraense, 4: 699-756.
- GORHAM, J. R., 1973, The Paraguayan Chaco and its rainfall, pp. 39-60. In: J. R. Gorham (ed.), Paraguay: ecological essays Academy of the Arts and Sciences of the America. Miami.
- GUIX, J. C. C., SALVATTI, M., PERONI, M. A. & LIMA-VERDE, J. S., 1989, Observations additionelles sur le comportement et la reproduction de Chelonoidis carbonaria (Spix, 1824) en captivité (Testudines – Testudinidae). Grupo de Estudos Ecológicos (Ribeirão Preto), Série Documentos, 1: 31p. (mimeo).
- HAIRSTON, C. & BUKCHFIELD, P. M., 1989, Management and reproduction of the Galapagos tortoise Geochelone elephantopus at the Gladys Porter Zoo. Int. Zoo. Yearb., 28: 70-77.
- HARLESS, M. & MORLOCK, H. (eds.), 1979, Turtle perspectives and research New York, John Wiley & Sons, xiv + 695p.
- HOUTMAN, H. & FOSSE, A. de la, 1989, Ervaringen met de verzorging en voortplanting van de kolenbrandershildpad (Geochelone carbonaria). Lacerta, 47(2): 40-46.
- IHERING, H. von, 1904, O Rio Juruá. Rev. Mus. Paulista, 6: 386-460.
- INSTITUTO NACIONAL DE METEOROLOGIA (Ministério da Agricultura), 1979, Normais climatológicas 2a edição. Rio de Janeiro, 78p.
- IVERSON, J. B. & EWERT, M. A., 1991, Physical characteristics of reptilian eggs and a comparison with avian eggs, pp. 87-100. In: D. C. Deeming & M. W. J. Fergusson (eds.), Egg incubation: its effects on embryonic development in birds and reptiles Cambridge University Press, Cambridge.
- LEGLER, J. M., 1963, Tortoises (Geochelone carbonaria) in Panama: distribution and variation. Amer. Midl. Nat., 70 (2): 490-503.
- MALLMANN, M. T. O., 1994, Influência da temperatura de incubação na determinação sexual em Geochelone carbonaria (Spix, 1824) (Reptilia, Testudines, Testudinidae) Dissertação de Mestrado, Pontifícia Universidade Católica do Rio Grande do Sul, 51p. (unpublished).
- MARITZ, M. & DOUGLAS, R. M., 1994, Shape quantization and the estimation of volume and surface area of reptile eggs. J. Herp., 28(3): 281-291.
- McKEOWN, S., JUVIK, J. O. & MEIER, D. E., 1982, Observations on the reproductive biology of the land tortoises Geochelone emys and Geochelone yniphora in the Honolulu Zoo. Zoo. Biol., 1: 223-235.
- MEDEM, F., 1960, Datos zoo-geograficos y ecologicos sobre los Crocodylia & Testudinata de los rios Amazonas, Putumayo y Caquetá. Caldasia, Bogotá, 8: 341-351.
- MEDEM, F., CASTAÑO-M, O. V. & LUGO-R., M., 1979, Contribución al conocimiento sobre la reproducción y el crecimiento de los "morrocoyes" (Geochelone carbonaria y G. denticulata; Testudines, Testudinidae). Caldasia, Bogotá, 12(59): 497-511.
- MOLL, E. O., 1979, Reproductive cycles and adaptation, pp. 305-331. In: M. Harless & H. Morlock (eds.), Turtles, perspectives and research John Wiley & Sons, New York.
- MOREIRA, G., 1989, Sympatry of the turtles Geochelone carbonaria and G. denticulata in the Rio Uatumã basin, central Amazonia. J. Herp., 23(2): 183-185.
- MOREIRA, G., 1991, Observações sobre Geochelone denticulata (Linnaeus, 1766) e Geochelone carbonaria (Spix, 1824) na bacia do Rio Uatumã, Amazonia central. Bol. Mus. Para. Emilio Goeldi (Zool.), 7(2): 183-188.
- MOSKOVITS, D. K., 1985, The behavior and ecology of the two Amazonian tortoises, Geochelone carbonaria and Geochelone denticulata, in northwestern Brasil Ph.D. Dissertation, University of Chicago. xviii + 328 p. (unpublished).
- PRESTON, F. W., 1953, The shape of birds'eggs. Auk, 70: 160-182.
- PRITCHARD, P. C. H. & TREBBAU, P., 1984, The turtles of Venezuela Soc. Study Amph. Rept., viii + 403p., pls.
- ROCHA, F. D., 1911, Chelonios do Ceará. Bol. Mus. Rocha, Fortaleza (2): 38-42.
- ROCHA, M. B., MOLINA, F. B. & CORREA, M. L. C. A., 1988, O comportamento de nidificação de Geochelone carbonaria (Spix, 1824), jabuti, em cativeiro (Testudines, Testudinidae). Resumos VI Encontro Anual de Etologia, Florianópolis: R15.
- ROSE, F. L., SIMPSON, T. R. & MANNING, R. W., 1996, Measured and predictged egg volume of Pseudemys texana with comments on turtle egg shape. J. Herp., 30 (3): 433-435.
- SNEDIGAR, R. & ROKOSKI, E. J., 1950, Courtship and egg laying of captive Testudo denticulata Copeia, 1950(1): 46-49.
- TASTEVIN, C., 1923, Nomes de plantas e animais em língua tupi. Rev. Mus. Paulista, 13: 687-763.
- TONGE, S., 1988, An analysis of the reproduction of a pair of red-footed tortoises (Geochelone carbonaria) over a twenty-year period. Dodo, 25: 82-90.
- UNDERWOOD, G., 1962, Reptiles of the Eastern Caribbean. Caribbean Affairs (N.S.), 1: 192p.
- VANZOLINI, P. E., 1977, An annotated bibliography of the land and fresh-water reptiles of South America (1758-1975). S. Paulo: Museu de Zoologia da Universidade de São Paulo. Vol. I (1758-1900), iv + 186p.
- VANZOLINI, P. E., 1996, A contribuição zoológica dos primeiros naturalistas viajantes no Brasil, pp. 90-238. In: A. M. Belluzzo (ed.), Dossiê Viajantes do Brasil, Revista USP, 30.
- VANZOLINI, P. E., 1997, A note on the reproduction of Trachemys dorbigni (Testudines, Emydidae). Rev. Brasil. Biol., 57(2): 165-175.
- VOKINS, A. M. A., 1977, Breeding the red-footed tortoise Geochelone carbonaria (Spix, 1824). Dodo, 14: 73-80.
- WILLIAMS, E. E., 1960, Two species of tortoises in northern South America. Breviora, 120: 13p.
Publication Dates
-
Publication in this collection
05 Apr 2001 -
Date of issue
Nov 1999
History
-
Accepted
26 Sept 1998 -
Received
10 Aug 1998